INTRODUCTION
In recent years, as nanotechnology has rapidly developed, nanomaterials have been widely used in the fields of biomedicine, pharmaceutical, and other industry. Of the various manufactured nanomaterials, silicon dioxide (SiO2) nanoparticles have the potential for widespread applications. SiO2 nanoparticles are being used in these fields such as chemical mechanical polishing and as additives to drugs, cosmetics, printer toners, varnishes, and food [1,2]. In recent years, the use of SiO2 nanoparticles has been extended to biomedical and biotechnological fields, such as biosensors for simultaneous assay of glucose, lactate, L-glutamate, and hypoxanthine levels in rat striatum [3], biomarkers for leukemia [4,5], DNA delivery [6,7], targeted drug delivery [8] and controlled drug release for genes and proteins [9,10]
Considering their wide range of applications, the potential adverse effect of SiO2 nanoparticles on human health and on the environment is of great interest. In this study, we investigated the potential harmful effect of exposure to SiO2 nanoparticles by conducting an in vitro toxicity assay with a focus on the involvement of oxidative stress. Numerous previous studies on nanoparticle toxicity, with various cell types and various nanoparticle types, reported that oxidative stress is one of the most important toxicity mechanisms related to exposure to nanoparticles [11-15]. Indeed, previous studies reported oxidative stress as the toxic mechanism of SiO2 [13,16-18]
In this study, to understand the potential harmful effect of nanoparticles on human health, the oxidative stress-related toxicity was investigated by exposure to SiO2 nanoparticles. SiO2-induced oxidative stress was assessed by examining formation of reactive oxygen species (ROS), the induction of superoxide dismutase (SOD) and heme oxygenase-1 (HO-1), as well as cytotoxicity effect was evaluation by cell viability. Subsequently, to understand the molecular mechanism of nanoparticle-induced oxidative stress, the involvement of oxidative stress-responding transcription factors, such as, nuclear factor-kappaB (NF-κB) and nuclear factor-E2-related factor-2 (Nrf-2), and mitogen-activated protein (MAP) kinase signal transduction pathway was also investigated.
MATERIALS AND METHODS
I. Cell Culture and Nanoparticle Treatment
Human bronchial epithelial cells, Beas-2B, were maintained in DMEM / F12 (GIBCO BRL Life Technologies, Rockville, MD, USA), supplemented with 10% (v/v) fetal bovine serum and 1% antibiotics, at 37℃ in a humidified atmosphere of air and 5% CO2. SiO2 nanoparticles were purchased from Sigma (St. Louis, MO, USA). Test solution of SiO2 nanoparticles were prepared in the culture media and dispersed for 20 minutes using a sonicator (Branson Inc., Danbury, CT, USA) to prevent aggregation. During the testing periods, the suspension of nanoparticles was stable and uniform in the culture media. For surface area measurement, Branauer, Emmett & Teller (BET) method was used with BELSORP-mini II, a volumetric adsorption apparatus (BEL Japan, Inc., Osaka, Japan). To investigate the size and shape of SiO2 nanoparticles, 20 uL of particle suspension form test media was dried on a 400 mesh carbon-coated copper grid and imaged with a JEM 1010 transmission electron microscopy (TEM; JEOL, Tokyo, Japan) at 40-100 kV. The size distribution of nanoparticles was evaluated using a Photal dynamic light scattering (DLS) spectrometer, DLS-7000 (Otsuka Electronics Co., Inc., Osaka, Japan). The concentration we used in this study was 1 mg/L to prevent aggregation and/or precipitation of particles. The cells were treated with various sizes of nanoparticles for 24 hours for toxicological studies.
II. Cell Viability Assay
Beas-2B cells were plated on 96-well plates and the plates were incubated with the nanoparticles for 24 hours. Cell viability was measured using 3-[4, 5-dimethylthiazol-2-yl]-2, 5-di phenyltera zolium bromide (MTT) reagent [19].
III. Flow Cytometry
Flow cytometry was performed on the treated and control cells for analysis of the cell cycle and apoptosis. Propidium iodide (PI) stained cells were analyzed using a flow cytometer (BD Science, San Jose, CA, USA) [20]. The effect on apoptosis was determined by the increase in the proportion of subG1 hypo-diploid cells.
IV. Formation of ROS
To detect ROS generation in nanoparticles-treated cells, a fluorometric assay using intracellular oxidation of 2,7-dichlorofluoroscein diacetate (DCFH-DA, Sigma) was performed [21,22]. Cells grown to confluence were treated with SiO2 nanoparticles for 24 hours, washed with PBS, and then incubated with DCFH-DA (40 uM) for 30 minutes. Following DCFH-DA incubation, the fluorescence of dichlorofluoroscein (DCF), which is the oxidized product of DCFH-DA, was visualized using a fluorescent microscope (Nikon, Tokyo, Japan) with excitation and emission wavelengths of 485 and 530 nm, respectively.
V. Western Blotting
For western blotting analysis, aliquots of the cell lysates were subjected to electrophoresis on a 12% sodium dodecyl sulfate (SDS)-polyacrylamide gel, and the gel transferred onto a nitrocellulose membrane (Bio-Rad, Hercules, CA, USA). The membranes were incubated with a primary antibody for 1 hours, subjected to further incubation with a secondary antibody and then exposed to X-ray film. Protein bands were detected using an enhanced chemiluminescence western blotting detection kit (Amersham, Little Chalfont, Buckinghamshire, UK). The antibodies for the detection of p38, phosphorylated p38, ERK-2, phosphorylated ERK and Nrf-2 were purchased from Santa Cruz (Santa Cruz, CA, USA), antibodies for JNK, phosphorylated JNK were from Cell Signaling (Beverly, MA, USA), antibody for HO-1 was from Stressgen (Victorya, BC, Canada) and antibody for Cu/Zn SOD was from Biodesign (Saco, ME, USA).
RESULTS
I. Characterization of SiO2 Nanoparticles
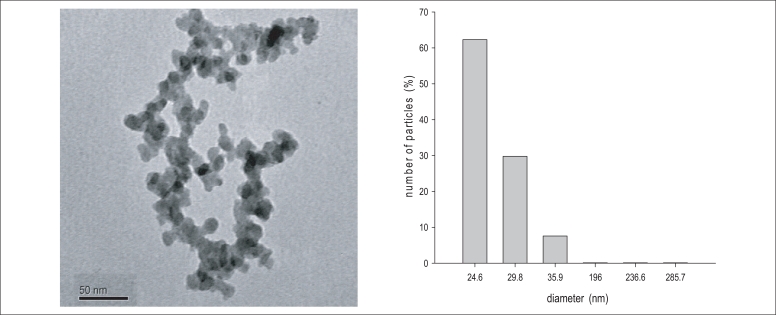
The result from characterization of SiO2 nanoparticles was summarized in Figure 1. The BET surface areas of was 201.01 m3/g, respectively. The TEM images indicated that silica nanoparticles had an even distribution with 20-40 nm individual particle sizes. The size distribution in the test medium was investigated using a DLS method, as the size of the nanoparticles distributed in the test medium were about 25 and 40 nm.
II. Cytotoxicity of SiO2 Nanoparticles
To investigate of SiO2 nanoparticle induced cytotoxicity effects on Beas-2B cells, cell viability and apoptosis were examined. MTT assay results show that decrease in cell viability was observed only about 20% compared with that of the control by SiO2 exposure (Figure 2). Flow cytometry analysis was conducted using PI staining to quantify the number of cells with a subdiploid DNA content (Figure 3). In Beas2B cells exposed to SiO2 nanoparticles, the number of cells in the subG1 phase increased. The degree of increase induced by SiO2 nanoparticles was about 3-fold compared with that of the control.
III. SiO2 Nanoparticles Induced Oxidative Stress
The formation of ROS was visualized in Beas2-B cells exposed to SiO2 nanoparticles by inspection under fluorescent microscope (Figure 4). DCFH-DA staining revealed increased concentration of ROS in SiO2 nanoparticles treated cells. As the increase in ROS formation was observed in SiO2 nanoparticle treated cells (Figure 4), a marker for cellular defense mechanism against oxidative stress (i.e. induction of antioxidant enzymes) was investigated in Beas2-B cells exposed to SiO2 nanoparticles (Figure 5). The expression of SOD protein was not changed, whereas that of HO-1 was increased by SiO2 nanoparticle exposure.
IV. Oxidative Signaling Pathway
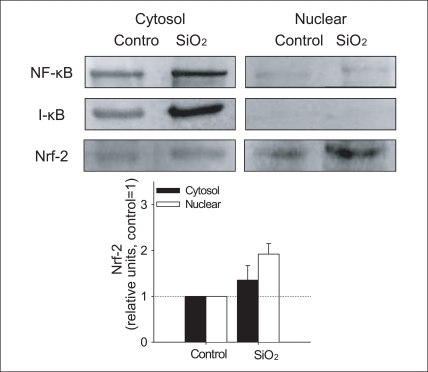
SiO2 increased formation of ROS and the induction of antioxidant enzyme, HO-1 (Figure 4,5). In order to understand the molecular mechanism of the observed oxidative stress and related physiological alteration by SiO2 nanoparticles exposure, NF-κB and Nrf-2 were examined in the cytosolic and nuclear fraction of cells treated with SiO2 nanoparticles. Neither nuclear localization of NF-κB nor the degradation of cytosolic I-κB was observed, whereas nuclear localization of Nrf-2 increased in cells treated with SiO2 nanoparticles (Figure 6). In our study, SiO2 nanoparticles induced the translocation of Nrf-2 into the nucleus as evidenced by the results of the western blot analysis (Figure 6).
To further clarify the possible upstream oxidative signaling pathway involved in Nrf-2 activation leading to HO-1 induction, we examined the activation of MAP kinases, known as major signaling kinases, involved in cell survival against oxidative stress through the Nrf-2 signaling pathway (Figure 7). The expression of unphosphorlyated forms of MAP kinases was constant regardless of exposure of SiO2 nanoparticles, whereas, the expression of phosphorylated form of ERK was strongly induced by SiO2 nanoparticles. The phosphorylation of ERK was significant by SiO2 nanoparticles (about 5-fold compared with that of the control). The expression level of phosphorylated p38 or JNK was not changed by SiO2 nanoparticles.
DISCUSSION
There are many studies have reported that nanoparticle toxicity deal with oxidative stress [11-14,23-27]. However, the mechanism by which oxidative stress is involved in nanotoxicity has been poorly addressed. We measured the ROS level (Figure 4), as well as the induction of antioxidant enzyme (Figure 5), provided strong evidence for the involvement of oxidative stress in SiO2 nanoparticles-induced cytotoxicity. To understand the molecular mechanism of the observed SiO2 nanoparticle-induced cytotoxicity by oxidative stress, activation of transcription factors (i.e NF-κB, Nrf-2) and signal transduction pathway (i.e. MAP kinase pathway) investigated.
In this study, to provide cellular consequence of the oxidative stress response observed (Figure 4-7), and cytotoxicity test were conducted (Figure 2,3). Exposed to SiO2 nanoparticles seemed to affect the cell viability and apoptosis by induction of ROS. ROS production was investigated as an initial step of oxidative stress, which revealed an increase in ROS formation in the cells exposed to SiO2 compared to control.
Neither nuclear localization of NF-κB nor the degradation of cytosolic I-κB was observed, whereas nuclear localization of Nrf-2 increased in cells treated with SiO2 nanoparticles (Figure 6). In our study, SiO2 nanoparticles induced the translocation of Nrf-2 into the nucleus as evidenced by the results of the western blot analysis (Figure 6). The translocation of Nrf-2 into the nucleus following nanoparticles treatment was associated with the increase in HO-1 protein, which suggests that nanoparticles activate Nrf-2 in association with the upregulation of HO-1 in Beas2-B cells. However, in this study, SiO2 nanoparticle-induced NF-κB activation was not observed, which was unexpected, as NF-κB is the major stress response transcription factor that has been reported to respond to a wide variety of environment stressors. Therefore, although SiO2-induced NF-κB activation was not observed in this study, the relative importance of NF-κB vs Nrf-2 signaling in terms of contribution to antioxidant response, such as HO-1 upregulation, may merit further investigation with various oxidative stress-inducing nanoparticles. To confirm SiO2 nanoparticles-induced for activation of transcription factor, more direct evidence, using electrophoretic mobility shift assay (EMSA) may be needed.
The upstream signaling mechanism responsible for regulating oxidative stress is poorly defined. Most studies have focused that oxidative stress may evoke the induction of antioxidant-related transcription factors, such as Nrf-2 or NF-kB via basal signal transduction systems, such as MAP kinase cascade. The MAP kinase cascades are multifunctional signaling pathways that are evolutionally well conserved in all eukaryotic cells. Three MAP kinase cascades that converge on ERKs, JNKs, and p38 MAP kinases have already been characterized [28,29]. Two of those three MAP kinase cascades converge on JNKs and p38 MAP kinases are preferentially activated by cytotoxic stresses, such as X-ray/UV irradiation, heat/osmotic shock, and oxidative/nitrosative stress [30-33]. To further clarify the possible upstream oxidative signaling pathway involved in Nrf-2 activation leading to HO-1 induction, we examined the activation of MAP kinases, known as major signaling kinases, involved in cell survival against oxidative stress through the Nrf-2 signaling pathway (Figure 7). The expression of unphosphorlyated forms of MAP kinases was constant regardless of exposure of SiO2 nanoparticles, whereas, the expression of phosphorylated form of ERK was strongly induced by SiO2 nanoparticles.
Activation of ERK MAP kinase pathway by SiO2nanoparticles suggests that the induction of HO-1 may be mediated through Nrf-2-ERK MAP kinase signaling pathway. ERK has been known to respond to internal stimuli, such as growth factors. However, our study revealed that SiO2 nanoparticles strongly induce the phosphorylation of ERK. Increase expression of phosphorylated ERK was also observed in our previous study with CeO2 nanoparticles exposure [34]. These results suggest that the ERK signaling pathway also responds to environmental stressors. The activation of the ERK signaling pathway by external stimuli has already been reported [30-33].
CONCLUSIONS
SiO2 nanoparticles exert their toxicity through oxidative stress as they cause the significant increase in cellular H2O2 concentrations. SiO2 nanoparticles induce induction of HO-1 via Nrf-2-ERK MAP kinase pathway. Our tested oxidative stress parameters are rather limited in terms of allowing the full understanding of oxidative stress and cellular response by SiO2 nanoparticle exposure. Further studies on the mechanism by which SiO2 nanoparticles induce the Nrf2-ERK MAP kinase pathway are warranted to better understand the nanoparticle-induced cytotoxicity by oxidative stress, as are studies with dose-response and time-course studies.